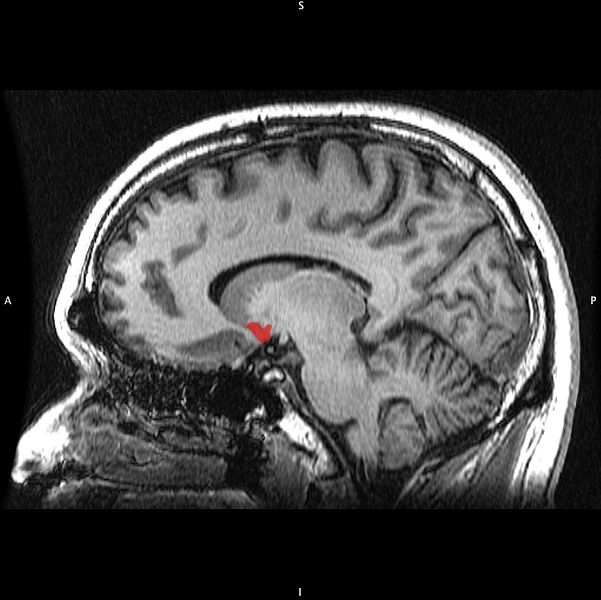
Sagittal MRI slice with highlighting (red) indicating the nucleus accumbens. The nucleus accumbens (NAc or NAcc), also known as the accumbens nucleus, or...